When reflected light enters the eye it is focused on an array of approximately 125 million photoreceptors in the retina. If the light is of the appropriate wavelength and intensity, it is absorbed by the photopigments contained in the photoreceptors and changes the electrical properties of the photoreceptors. These electrical signals are converted into chemical signals (neurotransmitters), initiating a cascade of signals through bipolar cells and retinal ganglion cells. Retinal ganglion cells send their axons through the optic nerve and up to the lateral geniculate nucleus (LGN). The LGN is a six-layered neural switchboard in the thalamus that routes the retinal signals to to primary visual cortex (V1), the first stage of visual processing in the neocortex. Cells in V1 respond to simple and complex features (oriented lines, colors, motion, binocularity) that give rise to perception. From V1, visual signals are further digested by area V2 and then parceled out to over 30 visual areas, each of whose functions are currently being investigated. Some areas have been shown to be especially important in the processing of motion signals (area MT); other areas are responsible for face perception (for example, area FFA); while other areas are responsible for depth perception (area V3a), or color (PIT). This is almost certainly an oversimplified schematic, but it is a heuristic that has helped shape experimental questions. Through critical reading of primary literature, it is possible to tease apart facts known about the visual system, and theories or hypotheses, which are important components of the scientific method (but still potentially false!).
Humans have two classes of photoreceptors, rods and cones. Rods are far more sensitive than cones, making them useful in low level lighting conditions, such as nighttime. Cones, being less sensitive, are used for daylight vision when more light is available. Cones are more concentrated at the center of gaze, the fovea, where no rods are found. The distribution pattern of the cones that parallels our visual acuity: highest at the center of gaze; lowest in the periphery. High acuity is critical to reading text, for example, which is why your eyes must continuously move across a line in order to read. Rods are found in highest concentration in a ring around the fovea, that peters off towards the periphery.
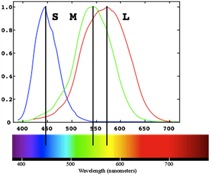
Most individuals have three cone types, referred to as long-, middle-, and short-wavelength sensitive (L, M, S) because they each respond optimally over a distinct range of the visible spectrum. It is the relative activation of these cone types that gives rise to color vision. An individual cone response varies only in the rate of firing, which depends on the wavelength of absorbed light and its intensity (the amount of light). This means that the same cone can respond in the same fashion to different wavelengths (say, long and middle) as long as the intensity is adjusted appropriately. This is known as the principal of univariance. As a result, a single cone response by itself cannot tell the brain anything about color. Color vision is achieved only in the presence of two or more cones with overlapping but different spectral sensitivity curves.
The photoreceptors talk to retinal ganglion cells via bipolar and horizontal cells. A given retinal ganglion cell listens to only a small group of photoreceptors, corresponding to a small region of the visual field. This is the cell’s receptive field. Different kinds of retinal ganglion cells have different physiological properties distinguished by receptive field size, color-opponency, direction selectivity, temporal and spatial frequency tuning, It is usually presumed that there are sufficient numbers of each class of retinal ganglion cell that the entire visual field can be tiled by a given class. This has led to the idea of “parallel” processing: the retinal image is sampled by multiple types of ganglion cells, each extracting a different “picture” of the visual world. Because of this, you can imagine vision as an amalgamation of several visual sub-systems over-layed on top of each other.
 Monet's painting
Monet's painting One useful system that attempts to connect neurobiology with psychology characterizes all extrastriate areas (areas downstream of V2) as areas concerned with “what” an object is versus areas concerned with “where” an object is located. The Where system is more primitive (evolutionarily older) than the What system, and is responsible for the perception of motion, space, position, three-dimensionality, figure/ground segregation, and the overall organization of the scene. This system is colorblind, has high contrast sensitivity, operates quickly, and has a slightly lower acuity than the “What” system. The “What” system provides us with the ability to identify objects and faces and to see in color. Because of the way these two pathways evolved separately, artists are able to create interesting effects by presenting different information to each system at the same time. In the example below Monet paints a rising sun in a color that has the same value (luminance to be precise) as the sky behind it, making it invisible to the Where system. Because the What system is still able to see it, the shape is somewhat unstable and appears to shimmer or jiggle.