Pharmacological Inhibition
The NCM (caudomedial nidopallium), a region in the avian brain analogous to Wernicke's area in humans, contains (part of) the neural substrate for the representation of tutor song memory in zebra finches (Bolhuis et al. 2006). Premotor regions in the avian brain such as HVC are also implicated in vocal learning; their role may be to encode sensory information used for successful imitation of tutor song (Roberts et al. 2012). Bilateral lesions to the NCM in adult zebra finches result in impaired tutor-song recognition, while leaving song production unaffected (Gobes and Bolhuis 2007). Taken together, these results provide evidence for distinct neural pathways for auditory recognition and vocal production of tutor song.
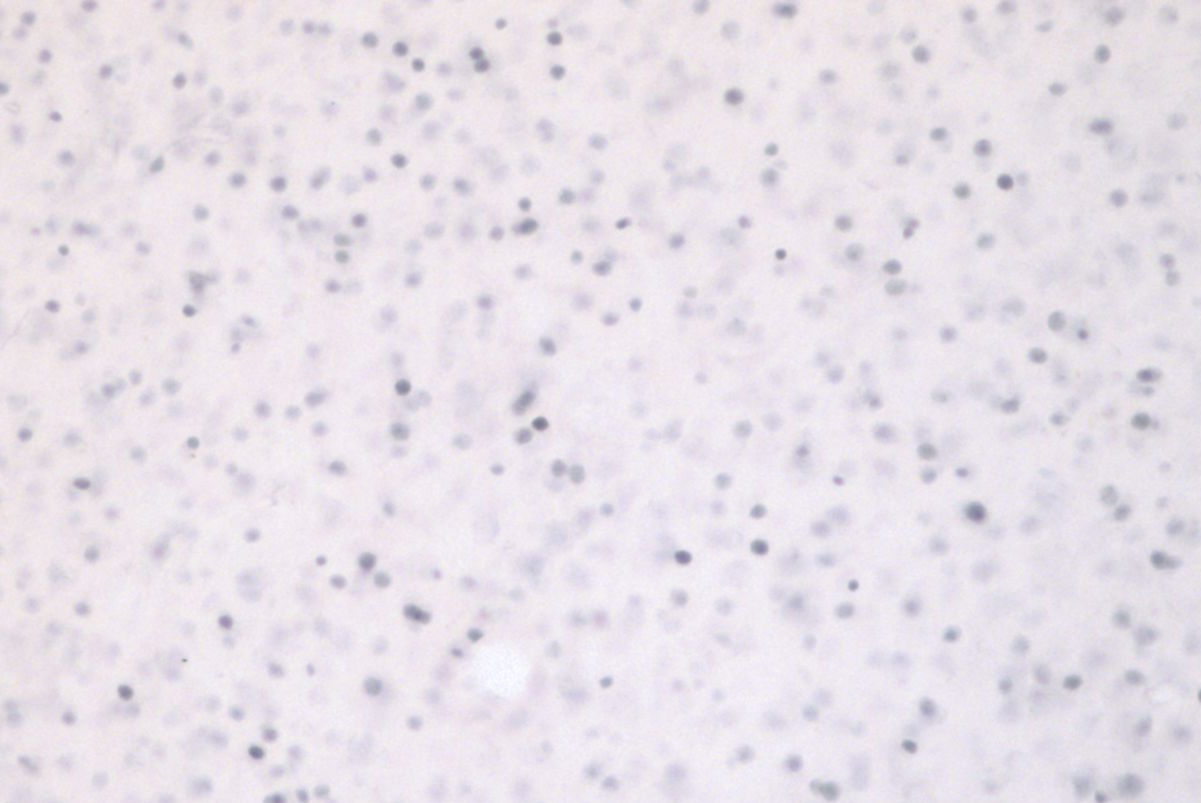
In our laboratory, we use Immediate Early Genes (IEGs), such as Zenk, as a measure for neuronal activation. IEGs are transcribed within 30min after stimulus exposure, and the protein products of these genes can be detected with Immunocytochemistry (Figure 1). The number of Zenk-expressing neurons in the NCM upon re-exposure to the tutor's song is related to the fidelity of song imitation, and is left-lateralized (Bolhuis et al. 2000, 2001; Moorman, Gobes et al. 2012). Hemispheric dominance of the left-NCM seems to develop with experience and is not innate (Moorman et al. 2012; Chirathivat et al. 2015). Interestingly, bilateral pharmacological inhibition of the molecular pathway responsible for inducing IEGs in the NCM during tutoring sessions impairs song learning in juvenile zebra finches (London and Clayton 2008). We are currently investigating whether development of a left-lateralized neural substrate for song memory is required for forming an accurate template, or memory, of tutor song with a combination of pharmacological and behavioral techniques.
References:
- Bolhuis JJ, Gahr M (2006) Neural mechanisms of birdsong memory. Nat Rev Neurosci 7:347–357.
- Bolhuis JJ, Hetebrij E, den Boer-Visser AM, de Groot JH, Zijlstra GGO (2001) Localized immediate early gene expression related to the strength of song learning in socially reared zebra finches. Eur J Neurosci 13:2165–2170.
- Bolhuis JJ, Zijlstra GGO, den Boer-Visser AM, Van Der Zee EA (2000) Localized neu- ronal activation in the zebra finch brain is related to the strength of song learning. Proc Natl Acad Sci USA 97:2282–2285.
- Chirathivat N, Raja S. C, Gobes S. M. H. (2015) Hemispheric dominance underlying the neural substrate for learned vocalizations develops with experience. Sci Rep 5: 11359.
- Gobes S. M. H. & Bolhuis J. J. Birdsong memory (2007) A neural dissociation between song recognition and production. Curr. Biol. 17:789–793.
- Gobes, S. M. H., Zandbergen, M. A. & Bolhuis, J. J. (2010). Memory in the making: localized brain activation related to song learning in young songbirds. Proc. R. Soc. B-Biol. Sci: 277, 3343–3351.
- London SE, Clayton DF (2008) Functional identification of sensory mechanisms required for developmental song learning. Nat Neurosci 11:579–586.
- Moorman S., Gobes S. M. H., Kuijpers M., Kerkhofs A., Zandbergen M. A., Bolhuis J. J. (2012) Human-like brain hemispheric dominance in birdsong learning. Proc. Natl. Acad. Sci. U.S.A. 109: 12782–12787.
- Roberts, T. F., Gobes, S. M. H., Murugan, M., Ölveczky, B. P., & Mooney, R. (2012). Motor circuits are required to encode a sensory model for imitative learning. Nature Neuroscience, 15(10), 1454–1459.