Color – Color in the Eye
Rods vs. Cones
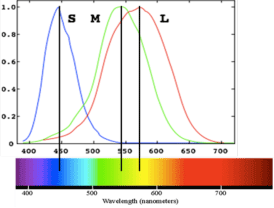
Humans have two classes of photoreceptors, rods and cones; we have one kind of rod and three kinds of cones, distinguished by differences in their peak spectral sensitivity. Rods are far more sensitive to visible light than cones, making them useful in low-level lighting conditions, such as nighttime. Cones are used for daylight vision when more light is available. A single type of photoreceptor, rod or cone, cannot by itself report color, which can be understood as a consequence of the principle of univariance, described below. To see in color, you need at least two different types of photoreceptors to be operating at the same time. Because we have three types of cones, we can therefore see color when cones are active—in other words during daylight. Did you know that in dim light, a moonless night, you cannot see color? Because the visual system processes information via parallel pathways, a loss of color vision at night does not constitute a loss of vision altogether. The ability to perceive forms and depth persists.
The Principle of Univariance
Most individuals are “trichromatsâ€, having three cone types, with peak sensitivity to long-, middle-, and short-wavelengths (L, M, S). An individual cone response varies in the degree to which it is activated, which depends on two variables:
- the wavelength of absorbed light; and
- the intensity of the light (the amount of light).
Wavelengths that match the spectral sensitivity of the cone type will be absorbed more readily and cause more activation of the cone; more intense light will also be absorbed more readily and cause more activation. The same cone could therefore respond in the same fashion to different wavelengths (say, long and middle) as long as the intensity is adjusted appropriately. This is known as the principle of univariance. As a result, a single cone response by itself cannot tell the brain the color of the stimulus. Color vision is achieved only in the presence of two or more cones with overlapping but different spectral sensitivity curves.
Any physical stimulus that elicits the same activity from the cones will appear the same. Because of the broad absorption curves of the cones, there are many physical stimuli (with different spectral distributions) that will elicit the same cone activity. These are referred to as metamers. The best example of metamers is yellow: consider monochromatic yellow—the region of the spectrum that appears yellow. This is physically entirely different than a mixed yellow made by mixing red and green light, yet the two yellows could be indistinguishable to normal trichromats.
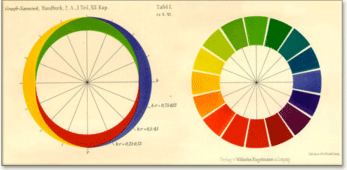
Color- opponency
Color-opponency describes the observation originally made by Ewald Hering, that certain pairs of colors appear to be mutually exclusive and cannot be mixed. For example, with a few very peculiar examples, colors cannot be described as “reddish greenâ€, or “bluish yellow†or “blackish whiteâ€. Opponency likely comes about because of the way in which retinal ganglion cells sample the cone signals.
The photoreceptors talk to retinal ganglion cells via bipolar and horizontal cells. A given retinal ganglion cell listens to only a small group of photoreceptors, corresponding to a small region of the visual field. This is the cell’s receptive field. Different kinds of retinal ganglion cells have different physiological properties distinguished by receptive field size, color-opponency, direction selectivity, temporal and spatial frequency tuning. The ganglion cells responsible for color-opponent processes can be divided into two classes: magnocellular and parvocellular. The Parvocellular cells deal with color information and can be divided into two groups: those that evaluate differences in firing of L versus M cones and those that evaluate differences between S cones and a combined signal from L an M cones. These cells were once thought to underlie Hering’s opponent colors, but careful analysis of the spectral tuning of LGN cells shows that this is not the case. But it is still widely assumed (and probably true!) that these cone-opponent cells are the building blocks for color computations performed by downstream areas.
The diagram below shows how the amount of red-or-green and yellow-or-blue combine to give rise to all hues: